Astrobiology of Comets
Richard B. Hoovera[1], Elena V. Pikutaa, Nalin C. Wickramasingheb, Max K. Wallisb, Robert B. Sheldona
aAstrobiology Laboratory, NASA/MSFC/NSSTC, 320 Sparkman Dr., Huntsville, AL USA 35805;
bCardiff Centre for Astrobiology, Cardiff University, UK.
ABSTRACT
We review the current state of knowledge concerning microbial extremophiles and comets and the potential significance of comets to Astrobiology. We model the thermal history of a cometary body, regarded as an assemblage of boulders, dust, ices and organics, as it approaches a perihelion distance of ~ 1AU. The transfer of incident energy from sunlight into the interior leads to the melting of near surface ices, some under stable porous crust, providing possible habitats for a wide range of microorganisms. We provide data concerning new evidence for indigenous microfossils in CI meteorites, which may be the remains of extinct cometary cores. We discuss the dominant microbial communities of polar sea-ice, Antarctic ice sheet, and cryoconite environments as possible analogs for microbial ecosystems that may grow in sub-crustal pools or in ice/water films in comets.
Keywords: comets, cryoconite, extremophiles, Orgueil, carbonaceous chondrites, meteorites, cyanobacteria
1. INTRODUCTION
Comets present the most interesting and important bodies of the solar system. Recent discoveries concerning microbial extremophiles, the chemical composition and thermal properties of cometary nuclei, and evidence for indigenous microfossils in CI carbonaceous chondrites suggest that comets may have far greater significance to Astrobiology than previously recognized. The role that cometary impacts played in the early delivery of the water crucial for the formation of the Earth’s atmosphere and oceans is now widely recognized and their role in the delivery of carbon, organics and prebiotic chemicals necessary for life is becoming better understood. An international armada of spacecraft from Russia, (Vega 1 and Vega 2), Europe (Giotto) and Japan: (Suisei and Sakigake) converged on Comet P/Halley in March 1986 and provided a wealth of new data about the nature, characteristics, and chemical composition of the nucleus. The observations established that the dayside surface temperature of Comet P/Halley is far above the 200 K sublimation temperature of ice. They also revealed that the albedo and composition of the nucleus of the comet has distinct similarities to the CI carbonaceous chondrites. These discoveries strengthen the case that the cores of extinct comets represent the most probable parent bodies for this important group of carbonaceous meteorites. Studies of a variety of extremophilic archaea, sulfur- and sulfate-reducing bacteria, cyanobacteria and diatoms have established that many representatives of these groups have astonishing resistance to UV, X-ray, g-ray and nuclear radiation from the decay of radioactive nuclei. These same groups also possess the capability to withstand repeated freeze-thaw cycles and exhibit long-term survival in ice and permafrost and during lyophilization conditions at low temperature and hard vacuum. Studies of cryoconite microbial ecosystems have provided new data concerning the microbial compositions of communities that thrive in water films surrounding rocks embedded in the ice of glaciers and the polar ice caps. These microbial consortia that can survive long periods of freezing, and then exhibit rapid growth during brief periods of elevated temperature and light levels in water films and cryoconite pools, are ideal models for the types of microbiota that might be capable of thriving on comets. Evidence for indigenous microfossils detected in situ in freshly fractured interior surfaces of CI and CM carbonaceous chondrites indicates that microbial life existed on the parent body at some point in time. Since comets represent probable parent bodies for CI carbonaceous meteorites, the detection of evidence for indigenous microbiology in the Orgueil meteorite may be interpreted as consistent with the concept that comets may, from time to time, provide habitats for microbial life.
2. THE DISTRIBUTION AND COMPOSITION OF COMET NUCLEI
It is now well established that comets are complex assemblages that are primarily composed of water ice, with embedded rocks, dust, frozen volatiles, clathrate ices, and complex organic chemicals that periodically orbit in the inner solar system. Study of the aphelia positions of the group of long-period comets (>200 years periodicity) allowed Oort to deduce the existence of a large cloud of small icy bodies arranged in a spherical distribution with a mean radius of about 50,000 AU.1 The Oort Cloud is thought to be comprised of trillions of these randomly oriented small icy bodies serving as a vast reservoir for new comets. These icy bodies are dislodged by passing stars or chaotic processes and sent to visit the inner solar system as long period comets such as Hale-Bopp (P~4000 years) and Hyakatake (P~8000 years). In contrast, the group of short period comets (P<200 years) have elliptical (typically prograde) orbits that are near the ecliptic plane. These comets were probably induced by collisions within the Kuiper Belt (30-50 AU). It is likely that the vast numbers of comets in the Oort Cloud and Kuiper Belt did not originate in those zones, but rather accreted within the Giant Planet Zone (5 - 30 AU). Gravitational encounters with Jupiter and Saturn (although not close enough for them to be accreted) could have ejected them into the Oort Clouds, while encounters with Neptune and Uranus could have tossed small icy bodies into the Kuiper Belt. Observations of comets with Hubble and the European Southern Observatory Very Large Telescope (which recorded an image of P/Halley in March, 2003 when it was over 28 AU from the Sun) have revealed a great deal about the behavior of icy bodies in the distant regions of the solar system. Comets do not develop comae or tails until they come nearer to the Sun than about the orbit of Jupiter (~5 AU, sometimes referred to as the “snow line”) and their water ices begin to sublimate. The coma (a dense cloud of water vapor, carbon dioxide and neutral gases sublimed from the nucleus) is surrounded by a hydrogen cloud (>106 km diameter). As comets approach perihelion they develop a long, curved, dust tail (~107 km) comprised of dust particles driven off the nucleus by escaping gases and a straight plasma ion tail (sometimes with rays and streamers) formed by interactions with the solar wind can extend for several hundred million kilometers (Fig. 1a).
It is generally accepted that the volatiles of comet nuclei are primordial ices that condensed on carbonaceous and mineral dust grains in the proto-solar nebula (far away from the hot central region) during the formation of the solar system. The “dirty snowball” or icy conglomerate model first advanced in 1950 by Fred Whipple2,3 was the dominant paradigm concerning the nature of the comet nucleus. The Infrared Astronomical Satellite (IRAS) that was launched on January 25, 1983 discovered 6 new comets and provided important new information about the nuclei of comets. Sykes and Walker4 showed that dust trails observed by IRAS in the wakes of comets have a refractory to volatile ratio of the order of 3, indicating that cometary nuclei are far dustier than previously thought. Comets are much more like “icy mudballs” than the Whipple “dirty snowballs”. The nuclei of comets are typically small (~10-20 km diameter), but some comets are much larger. For example, the great comet of 1729 (Comet Sarabat) has a 100 km diameter5 and the outer solar system body 2060 Chiron is 250 km diameter. This large body may also be a comet as it has exhibited some cometary activity and is in an unstable orbit that may lead to an Earth crossing path within 1 million years. 6
Comets are much richer in water and volatiles than are the inner planets. Most of the surface is covered with an extremely dark semi-inert, black carbonaceous crust (Fig. 1c). Volatiles and dust are ejected in geyser-like jets from increasingly active parts of the surface as the comet approaches the Sun (Fig. 1d). Comet rotation periods have been inferred from ground-based observations to be upwards of a few hours. Halley's comet appears to be unusually slow with a complex rotation with precession and periods 2.2 and 7.2 days.7 In 2003, the Hubble telescope observed Comet 67P/Churyumov-Gerasimenko (target of ESA’s Rosetta mission) and found it to have a 3 km x 5 km ellipsoidal nucleus with a rotation rate of ~12 hours. Space probes found the rotation rate of the nucleus of Comet P/Halley to be 52 hrs (Giotto) to 53 hrs (Vega). Giotto detected surface features (craters, ridges, mountains, etc.) and emitting vents that were found to be much more active on the sunlit side than on the night side of Halley’s nucleus (Fig. 2a). Dust grains of possible chondritic composition, and organic “CHON” particles were detected.
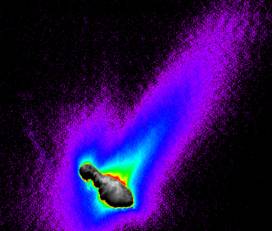
In 2001 the Deep Space 1 probe obtained a similar result for Comet 19P/Borrelly (Fig. 1d) which, like Halley, has a strongly elongated shape with a highly variegated, extremely dark, 8 km long nucleus. The rotation period of the nucleus is still being refined but is currently thought to be ~ 24 hours. The Deep Space 1 Short Wavelength Infrared Imaging Spectrometer (SWIR) found the nucleus of Comet 19P/Borrelly at 1.4 AU (perihelion 1.358 AU) to be very hot (from ≤300 K near the terminator to a maximum of ~ 340 K) and very dry (no trace of water ice detected).8 The nucleus of Comet 19P/Borrelly has the lowest albedo (~0.03) yet observed.
Comets contain a host of complex organic chemicals, macromolecules and polymers.9,10 Evidence continues to mount that comets played a crucial role in the origin of the atmospheres and the oceans of Earth and the introduction of carbon and organic/prebiotic chemicals to the early Earth during the period of heavy bombardment in the Hadean (4.5-3.8 Gyr).11 Water is an absolute requirement for all known life forms on Earth and it is now clear that comets are primarily water, but they are also rich in organic chemicals and complex molecular polymers. Hydrogen sulfide, hydrogen, sulfur and other potential sources of energy for a variety of microbial chemolithotrophs12 have also been detected.

a. b.

c. d.
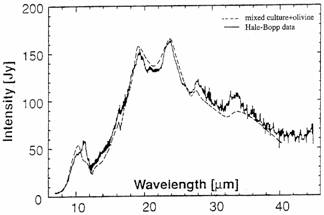
Fig. 1.a. Comet Hale-Bopp on April 5, 1997 (5 Days after Perihelion) and b. Infrared Spectrum of Comet Hale-Bopp (at 2.9 AU) compared with the spectrum of diatoms and olivine and Deep Space I images of comet P/Borrelly (Sept. 22, 2001) showing c. 10 km long cratered nucleus and d. orientation of jets of gas and dust.
Photos Courtesy: a. - University of Hawaii; c. & d. - NASA/JPL
In 1980, Hoyle and Wickramasinghe13 advanced the hypothesis that bacteria could be shielded from radiation and survive within comets and estimated that Earth could receive as much as 1018 cells per year from space. They have shown that the infrared spectrum of Comet Hale-Bopp is an excellent match for a mixed culture of bacteria, diatoms, and olivine14 over the 8-40 mm wavelength range. (Fig. 1b.) It has subsequently been shown that diatoms, cyanobacteria, and other ice microbiota have properties and characteristics ideally suited to surviving repeated freeze-thaw cycles. These microorganisms thrive in thin water films in permafrost and in cryoconite environments. This ability make them ideal candidates for life forms for not only survival, but also for carrying out active metabolism and growth on comets during the brief periods when temperatures are sufficiently high to allow liquid water, gases and microbial assemblages to collect around dark rocks embedded within voids in the rocks, or carbonaceous crust and enclosed cavities with the ice.15,16 Within the past few years, the role comets played in the distribution of pre-biotic chemicals and delivery of biochemicals critical to the origin of life on Earth (and intact viable microbes over the Cosmos) has become more seriously considered.17,18
Comets are considered to consist of pristine material from the formation of the solar system. However, as pointed out by Huebner19 and others, we must be cautious with the interpretation of the term “pristine.” The nucleus surface layers have been altered by exposure to UV and cosmic rays as well as hypervelocity dust impacts and decay of radionuclides in the nucleus throughout the 4.5 Gyr residence in the solar system. The interior water-ice may have been liquid for the first 0.01-0.1 Gyr due to radioactive heating via decay of 26Al and 60Fe and the release of energy during phase changes from amorphous to crystalline water ice may also contribute.24 Once a comet enters the inner solar system, it loses mass rapidly. ISO data revealed that water was the primary volatile (75-80 %) of the 40-50 km diameter nucleus of Comet Hale-Bopp. Minor fractions of CH4, NH3 and H2CO may come from clathrates (H2O ice with simple gases like CO2 and NH3 in a stable lattice) or result from atmospheric chemistry. ISO data obtained on September 27, 1996, indicated Hale-Bopp was releasing water vapor, carbon monoxide and carbon dioxide at a rate of 2 x 109 kg/sec. Volatiles and solids are well mixed throughout the nucleus of a new comet during its first approaches to the Sun. As comets lose ices they would develop an inert crust from the less volatile material.21
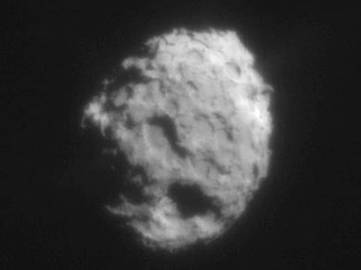
The first flyby of the nucleus of comet P/Halley, which is in a retrograde orbit, was achieved on March 6, 1986 by the Soviet spacecraft Vega 1. On March 9, Vega 2 passed within 8030 km of the nucleus. The Vega images showed a surprisingly dark (~ 14 km long) nucleus with two bright jets. Four days later, the Halley Multicolor Camera of the ESA Giotto spacecraft obtained spectacular images from 600 km showing the full contour of the nucleus (Fig. 2a). Recently, the Stardust probe at 500 km from comet Wild-2 (Fig. 2.b) likewise found a pattern of jets, including two apparently from the nightside.

a. b.
Fig. 2.a. Halley Multicolor Camera on the ESA Giotto spacecraft reveals the 16x8x8 km nucleus of Comet P/Halley, one of darkest objects in solar system, with bright geyser-like jets of escaping water vapor, gas, and dust. b. Nucleus of Comet P/Wild 2 from 500 km obtained by the NASA Stardust spacecraft during its closest approach on January 2, 2004.
(Photos Courtesy: a. ESA HMC and b. The Stardust Team, NASA/JPL).
The temperature of P/Halley’s nucleus20 was measured by the Vega 1 IKS infrared spectrometer at r = 0.8 AU and found to be in the range of 320-400 K (47 –127 oC). The IR observations also seemed to indicate that the hottest point was not subsolar. All the temperatures measured were considerably higher than the sublimation temperature of water ice in space (~200 K).23 These results were interpreted as an indirect confirmation of restricted activity of the nucleus and the existence of thermally insulating crust or mantle that covers large parts of the surface. They clearly establish that at a distance of 0.8 AU, the surface of Comet P/Halley was sufficiently hot to melt water ice. Confined within cavities and voids in ice or rocks or trapped in ice in contact with heated rock or black crustal material, gas pressures in excess of the critical pressure for liquid to form might result. This implies that true psychrophiles (which die when subjected to temperatures much above 0 oC) are not ideal candidates for life forms that might thrive (or even survive) on comets. Microorganisms capable of growing over a broad range of temperatures are better candidates. Many genera and species of small pennate diatoms and cyanobacteria satisfy this requirement. Cyanobacteria such as Mastigocladus laminosus, Calothrix spp. and Leptolyngbya spp. are well known thermopiles. Microcoleus chthonoplastes and several species of Phormidium inhabit permafrost and frozen soils as well as hot soils near thermal vents and fumaroles on the Antarctic volcano, Mt. Erebus. We have carried out theoretical modeling to examine conditions in which liquid water films could be present on comets.
3. THEORETICAL MODELS OF COMETS
Let us consider a comet made substantially of ice, with substantial admixtures of complex organics. Its surface is in part covered with a semi-inert carbonaceous crust and in part with sublimating ice.19 We consider the thermal behavior of solid ice-organic models, with differing degrees of porosity to investigate regimes of transient subsurface melting. Klinger22 gives the thermal penetration scales, using the thermal conductivity of crystalline ice:
k = ko / T where ko = 567 W/m for temperature T measured in degrees K.
The characteristic thermal skin depth for a comet rotating with a period of P ~ 2-10 hours is Ö(kP/prc) = 50-130 mm. Over orbital times of 1-3 years, the thermal scale extends to 5-10 m. A third distance scale is given by sublimation-erosion. With most incident energy going to latent heat, the radiation balance equation for an ice surface normal to the solar radiation at 1AU gives a sublimation rate of 7 x 1018 H2O-molecules/s (assuming a surface temperature Ts = 215 K).23 This amounts to 2 mm/hr (~4 meters/yr allowing a 4-fold reduction for varying orientation). Thus the surface retreats via sublimation at comparable speed to the penetration rate of the thermal wave.
Model 1: Translucent Ice (thickness z) over buried rock or absorptive layer of organic matter.
The incident radiation goes mainly into sublimation and reflection/re-radiation, but a fraction penetrates to be absorbed in the rock or organic layer and is then conducted upwards and downwards.
In the quasi-steady state, the 1-D conduction equation integrates as
T = Ts exp (z Fup / ko),
where T is the temperature at depth z and Ts is the surface temperature. The upward thermal flux Fup is a fraction of the radiation traversing upwards at the surface. If Fdn is the downward thermal flux and Fin is the absorbed incident radiation,
Fup + Fdn = Fin / (1+ 0.5t),
where t is the light scattering optical depth. We note that Fin is only a fraction of the solar constant Fo = 1.4 kW/m2 at 1 AU. We define the depth at which melting occurs marginally:
z* = ln(273/215) ko / Fup ~ [(1+ 0.5t)Fo / Fin] 0.2 m, (1)
where we have approximated Fdn @ Fup . Since the multiplying factor on the right-hand side of (1) is of order ~ 10, the melting of ice occurs at a depth of the order of meters.
If the absorbing rock or biomatter is deeper than z*, the ice melts above it up to depth z*, given sufficient time to reach the steady state. Hence, in practice, biomaterial could grow at the ice/water interfaces and the heat could be convected away by any motion of the water below it, enabling the ice to melt downwards.
Formula (1) shows the ice has to be translucent to meter-depths, for the optical depth t µ z.
Model 2: Crust with admixture of ice/frost
In an icy crust with pores up to a few mm in size, the thermal conductivity may be much lower24, k ~ 0.17 W/m oK for r = 0.25 g/cm3. The steady state thermal diffusion equation has a linear solution:
T = Ts + (Fup /k) z
with melting at a depth z* given by
z* ~ 2 x 58o (k/ Fo )[(1+ 0.5t) Fo /Fin ].
We can write this as z* = z(1 + 0.5 z*/l ) which solves as z* = 1/(1/z - 0.5/l). The condition for a solution is that z = 2 x 58o (k/Fin ) > 2l. If half the incident flux goes into the crust, then z = 3 cm and we have a solution with water at z* only if the radiation scattering path l exceeds 1.5 cm. This is an order of magnitude too high, so the mechanism appears to work only for higher solar flux as for a comet much nearer the sun. This conclusion is still only tentative as the vapor transport probably dominates the conductivity above 200 K in terrestrial snow23,25 and sublimation is competing with radiative/conductive heating in the few cm thick layer.
Model 3: Black, highly-conducting rocks, partially-submerged in ice
Because rock absorbs sunlight and is highly conducting, it can transport heat into the snow and maintain liquid water at the interface. Since thermal capacity is also higher for rock than for ice, the skin depth for diurnal rotation is a little larger (1.5-2 times) than in ice. Thermal conductivity for marble is k = 13 W/m oK (greater than silica and slate, but a third of that for quartz).
To estimate this for small rocks (radius X, smaller than skin depth 300mm for P=10 hours), consider the equation of flux balance at the interface:
k ¶T/¶x = const; krock (To – Tx ) / X @ kice(Tx – T1 ) / Skin(ice) (2)
Here To is the rock surface, 300-340 K in the midday sun at 1 AU, T1 is in the comet at a couple of skin depths, 150-180K. Since krock/kice @ 6 at 273 K, equation (2) formally gives an interface temperature Tx exceeding 279 K for X £ Skin(ice) and the entire T ranges given. Thus we expect either that heat is carried away by sublimation close to the outer surface if the ice is porous, or the ice to melt at deeper parts of the rock interface. On the diurnal cycle, the interface would cycle between the liquid and frozen states, as the rock is rather smaller than its thermal skin depth. Note that as the ice sublimes away, such rocks are left on the comet surface and would vaporize or melt the ice floor under them. Though the pool is shaded from the sun, scattered light in the ice is available for microbial photosynthesis. It is well known that photosynthetic cyanobacteria and bacteria thrive in mats on the floor of perennially ice-covered lakes in Antarctica.
Model 4: Kerogenous Crust over ice-liquid pool
Halley has a very dark crust which reaches temperatures up to ~400 K in full sunlight. The emissions from holes or cracks (~10% of surface) are confined to the dayside, implying rapid cooling and self-sealing, and that the crust is no thicker than a skin-depth. Halley is a strongly prolate body with complex rotation - its spin vector being inclined to its angular momentum vector. No part is in strong sunlight for over 20 hours though the precession period is 90 hours.26
The black crust of mainly organic (kerogen-like) matter has an outer layer that is porous and brittle (due to repeated daytime frying as well as micro-meteorite impacts), overlying a tougher asphalt-like layer whose original water has been replaced by organic gases diffusing from below and reacting chemically to seal most pores and cracks. Crustal thicknesses are determined therefore, by the peak solar heating and the chemico-thermal processing. Because of the self-sealing property, the underlying ice could melt periodically to form liquid pools, and with space above filled by vapor, because the original ice/snow mix is not compact. Such a vapor layer could inhibit inwards heat transfer, but latent heat transport (evaporation and re-freezing) and wave mixing (waves and sloshing due to the irregular precession) tend to bring the water temperature close to the 273 K of the lower ice surface. Without such mixing, a pool when heated from above might become thermally stratified (above a thin mixing layer near the lower ice due to the 4oC density inversion) permitting a rather warmer upper water layer.
The thermal conductivity k2 of the asphalt-like crust would be a factor 10 higher than the outer porous 'burnt toast' layer for which the value of Model 2, k1 ~ 0.17 W/m oK, can be taken. (Assume for simplicity a transition between the two at a temperature of 70 oC, for an outer temperature of 400 K). Adopting a thermal capacity for asphalt (rc=0.8 J/cm3K), the skin depth for P/p=10-20hr is 27-40 cm. The steady-state thermal diffusion equation as is appropriate to smaller thicknesses than these values again has linear solutions giving temperature differences across the outer toast and asphalt layers as:
DT1 = d1 (Fdown /k1) = 57oC DT2 = d2 (Fdown /k2) (3)
The condition for forming a watery pool is simply DT2 > 70oC or d2 / d1 < 70k2 / 57k1) » 12. Thus, within the uncertainties of our adopted parameters, this model shows that watery pools could form under crust composed of a ~10-20 cm asphalt layer with ~1-2 cm “burnt toast” exterior. The 10-20 cm thickness is plausibly sufficient for a nearly tight seal that prevents free evaporation and keeps the interior pressure well above water's critical pressure (~10 mb) and it is sufficiently smaller than the skin depth to cool quickly as the sun sets on Halley and to shut off gases leaking through the few cracks.
With these parameters, the downwards conductive heat flux Fdown = 57oC x (k1/d1) =0.5 - 1 kW/m2, i.e., 25-50% of the solar constant at 0.8 AU. In 10-20 hr this is sufficient to melt around 2 m of ice. Allowing for say 50% loss through leaks of vapor, it would seem that the depth of sub-crustal pools would fluctuate by 1 m via melting in the daytime and refreezing in the evening, night and early morning. The full depth of the pools depends of course on the orbital history, including particular orientations relative to the Sun. When observed by Vega post-perihelion (0.6 AU), the pools were probably past their maximum depth, but this requires specific modelling that includes thermal lag effects. The transition in the crust between the “burnt toast” exterior and the asphalt-like interior would also depend on history, so the above assumption of a transition at the 70 oC position at 0.8 AU needs refinement. The above models apply to a slow rotator like comet Halley. Comet Wild-2 presumably rotates faster, because it showed some jet activity from the nightside.
Model 5: Fluid instabilities on a rotating comet
Heat conduction in subcrustal H2O dominated material is heterogeneous, and depends upon the fluid or solid states. When ices melt, the porous nature of both the insulating mantle and underlying snow would be filled in, causing greater heat transport. Melting then appears to be a positive feedback mechanism. Aggravating this melt instability, we also suggest that the spin of a comet will enhance heat transport via promoting convection, particularly in equatorial regions, due to the Rayleigh-Taylor instability. When heated from above, fluids are generally stable to convection, but when cooled from above convective eddies promote more rapid heat transport and cooling losses. Thus for the slow-rotating comet (Model 4), nighttime thermal losses are enhanced compared with daytime thermal gains. But what of a fast-rotator with negative effective gravity?
That some comets may rotate fast enough to reverse gravity in an equatorial belt (and potentially cause the nucleus to split) has long been an issue. For this to occur, the period P has to be short enough 7 :
P = 2p / W < Ö(3p / rG) = 3.3 hr × s / Ör (4)
where s is a shape factor and r the mean density. For spherical bodies s = 1, whereas a right circular cylinder with a 2:1 aspect ratio (~ Halley) rotating about a short axis has s @ 1.7. If some comets have low density 0.3-0.4 g/cm3 and are spherical with P < 6 hr or elongated with P £ 10 hrs, this negative gravity situation could apply, though it appears marginal on current numbers.7 In this situation, an interior pool or lake is expected to be in contact with the crust while vapor occupies space further in. Convective mixing would operate under heating from the crust (daytime) but thermal stratification would occur under cooling (nighttime). Depletion of the regolith via spin-off would also occur in the same equatorial band, so any insulating dust layer is lost.
Under negative gravity in the equatorial belt of a fast rotating comet, the liquid pool rests on the crust with a vapor layer separating it from interior ices. The pool is heated from the crust in the daytime and cooled by evaporation into the vapor layer whose temperature decreases to the 0oC of the interior ice. Both the liquid and vapor layers are convectively unstable. But nighttime when the temperature gradients reverse, the opposite is the case, convection is suppressed and heat transport slowed. Thus the sub-crustal layers act to some extent as a heat pump.
The Rayleigh-Taylor instability that generates convective cells has a growth rate:
g = [g k (r1 - r2)/(r1 + r2)]1/2 (5)
where k is the reciprocal of the vertical gradient (1/length), and r1, r2 are the densities of two layers. Consider eg. a spherical comet with r=0.3g/cm3, P=4 hr and effective g ~ 0.1 cm/s2 (cf. the surface gravitational acceleration g of a 10 km iceball is 0.25 cm/s2). Convective cells in a 1 meter (=p/k) vapor space with DT=20oC have effective growth times from (5) of approximately 100 seconds. Fifteen minutes – i.e. nine e-folding times - would see convective cells fully developed. Thus convection is sufficiently fast to be effective. Similar convective cells in a 1-meter pool (20-40 oC; r1-r2 = 0.006) take several times longer to develop.
During the nighttime, as the crust becomes cold, the pool cools. In this case (pool cooled from “bottom”), convection is suppressed until the 4 oC temperature inversion of water is reached. When the interface drops to 0oC, the water density increases by ~ 1x10-4 and the convective instability growth time (for ~1 cm cells between the 0 oC and 4 oC isotherms) is ~ 200 s. As several e-folding times are needed, this may not be fast enough to cool the whole of the pool to 4 oC before freezing starts at the crust interface and progresses inwards. Thus we can conclude that the R-T instability and convection hardly operate in the nighttime cooling phase, implying that heat loss is relatively slow compared with heat gain from daytime insolation (if mixing due to waves and sloshing of Model 4 is suppressed). As well as this “heat pump” difference, the negative gravity configuration with pool in contact with the crust might give different behavior to a Halley-type slow rotator, e.g., more erratic, explosive releases through less frequent crustal cracks.
4. MICROBIOTA THAT MIGHT SURVIVE ON COMETS
Theoretical calculations as well as observations of the nucleus of comet P/Halley indicate that internal temperatures in localized regions within the comet can vary over an extremely wide range. As true psychrophiles perish when exposed to elevated temperatures, better candidate organisms are those forms that can survive, and even thrive over an extremely wide range of temperatures, salinity, pH, and concentrations of sulphur and sulfates. These include groups such as the diatoms (eukaryotic forms) and many cyanobacteria, crenarchaeota, and sulphur and iron bacteria and archaea. We review some relevant information concerning these groups:
Diatoms are dominant Eukaryotic life forms of the polar sea-ice
• Live in liquid water/ice interface (Temperature ~ 0 oC).
• Sympagic community forms in small brine-filled spaces between ice crystals in the 3-D matrix. Many polar diatoms live in sheaths and mucilaginous tubes (Berkeleya sp.);
• Amphora antarctica is dominant and Melosira arctica grows in brine channels and on ice floes.
• Transpolar data by American and Russian scientists drifting in ice camps in 1994 shows high activity in sea-ice, with over 200 species of diatoms and a very large community of other ice algae, ice bacteria, viruses and fungi are present.
• Diatom mat underneath sea ice in McMurdo Sound included 32 species. Amphiprora kjellmanii, A. oestrupii, Biddulphia weissflogii, Coscinodiscus subtilis, Eucampia balaustium, Fragilaria linearis, Nitzschia martiana, N. seriata, Pleurosigma antarcticum, Rhizosolenia alata and R. rostrata.
• Epicryotic diatom community under sea ice are distinct from plankton. Antarctic diatoms are grazed by protists (ciliates, radiolaria, foraminifera, flagellates, choanoflagellates) and zooplankton (copepods, krill).
Cyanobacteria are dominant life forms on the Antarctic ice sheet
• Cyanobacteria (blue green algae) are photoautotrophic bacteria.
• All species are aerobic—but a few species are facultative anaerobes: chemolithotrophs using H2S at high temperature.
• Contain chlorophyll A and photosynthetic pigments - phycoerythrin and phycocyanin (blue green, green, yellow, red, violet, dark blue and black) -- Red Sea named for Oscillatoria spp. blooms.
• Cell division by binary fission.
• Trichomes encased in mucilaginous sheaths for motility and protection from desiccation and radiation.
Cyanobacteria are the dominant life forms of the Antarctic continental desert.
• Growth and reproduction totally dependent on minute pockets of liquid water that have major fluctuations in volume, osmolarity, pH, and temperature during freezing and thawing.
• Must withstand extremes of temperature, salinity, alkalinity, dessication; frequent freeze-thaw cycles; intense UV radiation; and alternate periods of intense sunlight and total darkness ---
These are the most comet-like conditions to be found on Earth.
Cyanobacteria are the dominant life forms of the Antarctic cryoconite communities
• Dominant species: Phormidium frigidum; also found are species of: Microcoleus, Lyngbya, Nostoc and Diatoms, Green Algae; Bacteria, Fungi, Actinomycetes and Yeasts.
Microbiota of permanently cold polar marine sediments
• Sulfate-reducing obligate psychrophiles (e.g. Desulfofrigus sp., Desulfofaba sp., & Desulfotalea sp.) have been isolated from permanently cold Arctic marine sediments.
Microbiota of Arctic Swamps, Permafrost, and Cryopegs
• Diatoms, cyanobacteria, yeasts, fungi, actinomycetes, methanotrophs and secondary anaerobes such as methanogens (archaea) and sulfate-reducers (both psychtotolerants and psychrophiles) grow in thin films between ice and mineral grains of permafrost.
• Sulfate-reducers, acetogens, and methanogens grow in supercooled (-15 oC) water-brine lenses within the permafrost (Cryopegs).27
Microbiota of Antarctic streams, lakes, and epilithon
• Phormidium and closely related Oscillatoriacea.
• Cyanobacteria are microbial biomass dominants in Antarctic stream and lake mats and sediments.
• Streams in the Dry Valley freeze in February and then ablate to leave behind desiccated epilithon of long black cyanobacterial strands on dry streambeds--dominant in mats are Calothrix and Gloecapsa-sheaths protecting cells from dessication.
Cyanobacteria, sulfur- & sulfate-reducing bacteria and archaea are dominant microorganisms in:
• Antarctic fumaroles and geothermal habitats.
• Lyngbya species have been observed to grow as dark grey to blue-black felt mats on the hottest slopes (40-60 oC) of the Mt. Erebus volcano in Antarctica.
• Phormidium fragile and N2-fixing Mastigocladus laminosus form red-brown felts at 30-40 oC with Gleocapsa magma and Stigonema spp.
Antarctic continental ecosystems
• In hypersaline, hyperalkaline lakes species of archaea, cyanobacteria and halophilic bacteria (e.g., Dunaliella spp. and Microcoleus spp.) are the microbial biomass dominants in layers of hydrohalite (NaCl*2H2O), gypsum (CaSO4*2H2O), and mirabilite (Na2SO4*10H2O) beds of the McMurdo ice-shelf lakes.
• Some cyanobacteria and some phototrophic sulfur bacteria have sensitive photoreceptors and can conduct photosynthesis at low light levels deep beneath thick crusts of ice. The deep chlorophyll a maxima at 60 M depth beneath ice crust of Lake Vanda in regions of high Sulfur and H2S (Rhodopseudomonas lacustris).
The actual activity and significance of psychrophilic microbiota on our planet are not yet thoroughly understood as the methodology for the cultivation of obligate psychrophiles has only recently been developed. Diatoms, algae, cyanobacteria, yeasts, bacteria & archaea are phototrophs, lithotrophs or heterotrophs (able to utilize light, H2S, or pairs of amino acids or other organics) making them candidates for growing in benthic mats in cryoconite pools, in thin films around mineral grains, and in permafrost or brine channels & rock/ice interfaces in cold or hot regimes under a comet’s carbonaceous crust.
- COMETS, METEOR STREAMS, AND METEORITES
The association between comets and meteor streams is now well established. In 1866, Schiaperelli first connected the Perseid meteor stream with the orbit of comet P/Swift-Tuttle.28 Many well known meteor streams are clearly associated with the passage of the Earth through dust debris left by periodic comets as seen in Table I.
Table I. Meteor Streams Associated with Comets
|
Meteor Stream |
Recent Peak |
Associated Comet (Period Yrs.) |
|
Lyrids |
April 22 |
P/Thatcher 1861 I (450) |
|
Aquarids |
May 6 |
P/Halley 1986 (76) |
|
Perseids |
August 12 |
P/Swift-Tuttle 1862 III (105) |
|
Orionids |
October 21 |
P/Halley (76) |
|
Taurid Complex |
N (Oct. 12-Dec.2); S (Sept. 17- Nov 27) |
2P/Encke (3.3) |
|
Leonids |
November 19 |
P/Tempel-Tuttle (33) |
|
Geminids |
December 6-19 |
Asteroid Phaethon (1.43) |
Comets have albedos that are very similar to those of the CI carbonaceous chondrites. Orbital and dynamic studies have indicated that extinct comets may comprise over half of the population of all near Earth objects and that comets represent likely parent bodies for some carbonaceous meteorites.29 Recent investigations have also explored amino acid chemistry for a link between carbonaceous chondrites and asteroids and comets. Glavin30 compared the abundances (relative to glycine) of the amino acids alanine, b-alanine, a-amino-n-butyric acid (ABA) and a-aminoisobutyric acid (AIB) in the Orgueil and Ivuna CI chondrites and compared them with the abundances in the Murchison and Murray CM chondrites. He concluded that the Orgueil and Ivuna chondrites are probably of cometary origin but that the amino acid differences could indicate that the CM meteorites came from a different type of parent body. Engel et al. 31 and Ehrenfreund et al.32 have clearly shown that the amino acids of the carbonaceous meteorites are extraterrestrial and indigenous. These studies were carried out with the most sensitive HPLC methods available and amino acids were identified at concentrations as low as 8 ppb. Only eight of the 20 protein amino acids that are common in all life on Earth are present in the meteorites. Their stable isotope composition and the fact that these amino acids are mostly racemic (and the absence of such life critical amino acids such as Serine, Threonine, Phenylalanine, Arginine, Lysine, and Histidine) provides dramatic proof that the amino acids are indigenous and extraterrestrial and that the meteorites have not been subjected to recent contamination by bacteria, fungi and other microorganisms. If the meteorites had been recently contaminated by living bacteria, fungi, or pollen, the other 12 protein amino acids would certainly have been detected.
The elemental abundances of both comets and the CI carbonaceous chondrites have been found to be very close to solar abundances, with the curious exception of the Iron/Silicon ratio which is 25 % that of the Sun. (Comet P/Halley has an Iron content that is overabundant by a factor of 2 and its Silicon abundance is half that of the Sun). The relative abundance of the elements in comets as compared with the lithosphere of Earth and the Orgueil CI carbonaceous meteorite, the Sun, and Bacteria are shown in Table II. The strong similarity atomic percent of the life critical elements (C, H, O, N, P, S) in comets and bacteria is most intriguing.
TABLE II. Relative Element Abundances in Earth’s Lithosphere and Comets
|
Element |
Lithosphere* % |
Comets* % |
Orgueil* % |
Sun/Orgueil* |
Bacteria % |
Atom% Comets* |
Atom% Bacteria |
|
C |
0.037 |
11.37 |
3.22 |
9.90 |
6.4 |
19 |
7.8 |
|
H |
2.9 |
54.64 |
2.02 |
- |
63 |
48 |
63 |
|
O |
60.2 |
28.34 |
46.5 |
1.82 |
29 |
21 |
26 |
|
N |
0.006 |
1.32 |
0.318 |
40.6 |
1.4 |
1 |
1.8 |
|
P |
0.079 |
- |
0.093 |
1.02 |
0.12 |
<0.002 |
0.17 |
|
S |
0.023 |
0.71 |
5.41 |
1.37 |
0.06 |
1.7 |
0.06 |
|
Na |
2.5 |
0.10 |
0.498 |
1.07 |
- |
0.24 |
0.11 |
|
K |
0.89 |
- |
0.054 |
1.03 |
- |
0.005 |
0.07 |
|
Ca |
1.9 |
0.06 |
0.932 |
1.07 |
- |
0.15 |
0.04 |
|
Mg |
2.5 |
0.99 |
9.61 |
0.94 |
- |
- |
- |
|
Si |
20.5 |
1.83 |
10.68 |
0.99 |
- |
- |
- |
|
Fe |
1.8 |
0.52 |
18.43 |
0.92 |
- |
- |
- |
|
Ni |
0.0025 |
0.04 |
1.08 |
1.05 |
- |
- |
- |
|
Al |
6.2 |
0.07 |
0.85 |
1.02 |
- |
- |
- |
|
Total |
99.53 |
99.99 |
99.695 |
- |
99.98 |
- |
- |
Under the current paradigm, comets are devoid of liquid water and are entirely sterile. This is clearly reflected in the NRC Report detailing criteria for return from Astromaterials, which states the following:
“NRC Report on Small Body Sample-Return:
All samples returned from planetary satellites and small solar system bodies that must be contained should be treated as potentially hazardous until proven otherwise.
No sample containment and handling is warranted beyond what is needed for scientific purposes for:
· Moon, Io, new comets, Interstellar Dust Particles (IDP)* with a High Degree of Confidence
· Phobos, Deimos, Callisto, C-type asteroids, undifferentiated metamorphosed asteroids, differentiated asteroids, all other comets, IDP’s* with a Lesser Degree of Confidence
Strict containment and handling are warranted for:
· Europa, Ganymede, P-type asteroids, D-type asteroids, IDP’s*
· Sample return provisions for contained samples are the same as for Mars
*Depending on parent body and time of exposure to space environment.” 33
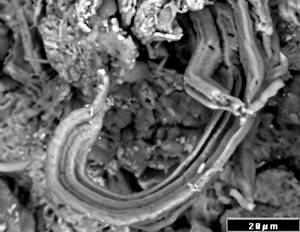
It is presently widely accepted that comets contain stores of water ice, and possibly played a significant role in the delivery of water to Earth billions of years ago helping form the oceans and atmosphere and making Earth suitable for life. On the “dirty snowball” model, the snow and ice go directly from the frozen state to water vapor (i.e. sublimate) in the very low pressures appropriate to space when the comet is within a few AU of the Sun. The scientific consensus is that water does not exist in liquid form on comets, and therefore comets cannot support life as we know it.33 Comets (Figs 2a and b), like asteroids (Fig. 3a & b), exhibit craters clearly establishing that their surfaces have a long history of impacts by the collisional debris on independent orbits about the Sun. The possibility that some of these impactors carry not only organics, but also intact, and possibly even viable microorganisms to comets or asteroids, cannot be ruled out. Some of the impacting debris would have originated like the Martian meteorites in occasional giant impacts that nevertheless do not extinguish microbial life. If impacts on Earth similarly eject material to space from polar ice sheets, sea-floor sediments, or crustal rocks, viable microorganisms might have transferred to comets and asteroids. The Orgueil CI meteorite, which may be a remnant of a comet, has been found to contain abundant microfossils of mineralized cyanobacteria (Fig. 3c & d) surrounded with high carbon (kerogen) sheaths.34 It is important to note that the ability of a small number of microorganisms to multiply by binary fission into phenomenally large numbers in short time periods when conditions are suitable have been well known to microbiologists.


a. b.

c. d.
Fig. 3. Galileo Image of a. Asteroid 243 Ida (Koronis Family) and b. the s-type Asteroid 951 GASPRA (20x12x11 km) 500 Mya. c. d. Images of indigenous microfossils34 exhibiting hollow filaments, trichomes and emerging cells of cyanobacteria in the Orgueil CI carbonaceous chondrite, which is of possible cometary origin.
6. CONCLUSIONS
It has been established that comets contain water ice, clathrate ices, complex organic chemicals, sulfur and hydrogen sulfide, and a suite of mineral constituents that are similar to those that are present in the carbonaceous chondrites. The international armada of spacecraft that converged on Comet P/Halley in March 1986 provided a wealth of new data concerning the nature of the nuclei of comets. They confirmed that the dominant component of the nucleus of Comet P/Halley was water ice. They also showed that the nucleus contained ices, complex organic chemicals, sulfur and hydrogen sulfide, and a suite of mineral constituents similar to those found in Carbonaceous Chondrites. The very black (albedo ~3%) carbonaceous crust covering the nucleus was an astonishing discovery. These results helped to clarify the profoundly important role that comets played in the origin of the oceans and the formation of the atmosphere of Hadean Earth. Their delivery of carbon, complex organic compounds and polymeric matter, and possibly even intact microbial components may have played a crucial role in the Origin of Life on Earth.35 Though comets are extremely cold and in dark conditions for most of their orbit, as they approach perihelion the sunlit surface of the nucleus is exposed to intense sunlight and can become very hot (~350 – 400 K). Our modelling analysis indicates that liquid water films and enclosed pools analogous to the cryoconite niches found around dark rocks embedded in glaciers and polar ice sheets could exist in some comets. Conditions are favorable for the production of evaporite minerals (epsomite, chlorite, gypsum, trona, etc.) as the water is lost from subcrustal pools by sublimation and evaporation. The 2004 flyby of comet Wild-2 showed depressions, aligned escarpments and tall pinnacles, indicating quite a thick crust and durable structures accreted from recondensing gases. These arguments and observations, combined with our study of indigenous bacterial microfossils found in-situ in the CI and CM carbonaceous meteorites (Fig. 3c and d) have firmed up the proposition that comets may harbor microbial life.
The information now available suggests that microbial candidates that might survive (or perhaps even thrive) on Comets must be able to withstand:
• Long periods of extreme cold, desiccation and anaerobic conditions
• Exposure to periodic high doses of UV, X-ray, gamma-ray and nuclear radiation
• Exposure to dramatic variations in pH, salinity and evaporite minerals
• Survival of spores or cells during very long periods in a frozen state
• Active and rapid metabolism and photoautotrophic, chemolithotrophic, or organotrophic growth during short periods and over a broad range of temperatures when sunlight, chemicals, etc. are available in water films (or entrained cryoconite pools) as the Comet approaches to within 1 AU of the Sun
These capabilities are found in few genera and species of Eucaryota: Bacillariophyceae (diatoms)
and in many genera and species of Procaryota from the Domains of Archaea and Bacteria:
Archaea: Crenarchaeota – Desulfurococcales & Sulfolobales; Euryarchaeota – Halobacteria & Thermococci
Bacteria: Aquificae, Thermotogae, Deinococci, Deferribacteres & Cyanobacteria
All the required ingredients seem to be present on comets to support and maintain assemblages and ecologies of many groups of extremophiles (psychrotolerants, thermophiles, halophiles, alkaliphiles, acidophiles, and radiation-resistant forms) within these microbial communities. Observations of P/Halley by Giotto and evidence of cratering of the nucleus of Comet 19P/Borrelly36 in Deep Space 1 images and of Comet Wild-2 as seen by the Stardust Mission37 clearly establish that these small icy bodies are covered with highly evolved crust rather than simple “pristine” solar system material. Recent discoveries have also revealed the ability of a number of microbial extremophiles to thrive in super-cooled hypersaline lakes that are perennially ice-covered, and in ice-bubbles, cryoconite voids and thin water films at brine ice/soil interfaces within permafrost (at temperatures as low as 220 K). These results undermine the paradigm that comets could not conceivably harbor life. Taken in conjunction with recently obtained evidence for indigenous microfossils in carbonaceous meteorites (of presumed cometary origin) we conclude that additional research and serious reconsideration is clearly warranted. Exciting new information about the nature, composition and potential for microbial life may be obtained when the Stardust samples are returned from comet Wild-2, when Europe’s cometary probe of the Rosetta mission touches down on comet 67P/Churyumov-Gerasimenko in 2014, and via collections of cometary dust from the upper stratosphere.
ACKNOWLEDGMENTS
We are grateful to the NASA/JSC Center for Biomarkers in Astromaterials for providing funding support for this research. We also thank Gregory Jerman and James Coston at NASA/MSFC for Electron Microscopy support and Academician Georgy Zavarzin, and Dr. Ludmila M. Gerasimenko of Institute of Microbiology, Russian Academy of Science for helpful discussions. We thank Dr. William Birch, Victoria Museum, Melbourne, Australia, (Murchison), Dr. Paul Sipiera, the Dupont Meteorite Collection and Planetary Studies Foundation (Orgueil, Bells, Acfer 324) and Dr. Claude Perron of the Musée Nationale d’Histoire Naturelle, Paris (Orgueil) for providing samples of carbonaceous chondrites used in this research.
REFERENCES
1. J. H. Oort, “The Structure of the Cloud of Comets Surrounding the Solar System, and a Hypothesis Concerning Its Origin.” Bull. Astron. Inst. Neth. 11, 91-110, 1950.
2. F. L. Whipple, “A comet model I. The acceleration of comet Enke.” Astrophys. J., 111, 375-394, 1950.
3. F. L. Whipple, “On the Structure of the Cometary Nucleus.” in The Moon, Meteorites, and Comets.” (B. M. Middlehurst and G. P. Kuiper, Eds.), University of Chicago Press, Chicago, 639-664, 1963.
4. M. V. Sykes, and R. G. Walker, “Cometary dust trails. I. Survey” Icarus, 95, 180-210, 1992.
5. D. Steel, “Cometary Impacts in the Biosphere” in Comets and the Origin and Evolution of Life. (Paul J. Thomas, Christopher F. Chyba, and Christopher P. McKay, Eds.) Springer-Verlag, New York. 216, 1997.
6. G. Hahn, and M. E. Bailey, “Rapid Dynamical Evolution of Giant Comet Chiron.” Nature, 348, 132-136, 1990.
7. D. C. Jewitt, "Cometary Rotation: An Overview", Earth, Moon and Planets 79, 35–53, 1999.
8. L. A. Soderblom, D. T. Britt, R. H. Brown, B. J. Buratti, R. L. Kirk, T. C. Owen and R. V. Yelle, ""Short-wavelength infrared (1.3-2.6 mm) observations of the nucleus of Comet 19P/Borrelly), Icarus, 167, 100-112, 2004.
9. A. Delsemme, “Nature and History of the Organic Compounds in Comets: An Astrophysical View” in Comets in the Post-Halley Era, Vols. I-II. (R. L. Newman , M. Neugebauer, and J. Rahe, Eds.) Dordrecht, Boston. 377-427, 1989.
10. W. F. Huebner, and D. C. Boice, “Polymers and Other Macromolecules in Comets.” in Comets and the Origin and Evolution of Life. (Paul J. Thomas, Christopher F. Chyba, and Christopher P. McKay, Eds.) Springer-Verlag, New York. 111-129, 1997.
11. A. Delsemme, “The Origin of the Atmosphere and of the Oceans” in Comets and the Origin and Evolution of Life. (Paul J. Thomas, Christopher F. Chyba, and Christopher P. McKay, Eds.) Springer-Verlag, New York. 54, 1997.
12. R. B. Hoover, and E. V. Pikuta, “Microorganisms on Comets, Europa, and the Polar Ice Caps of Mars” Instruments Methods and Missions for Astrobiology VII, SPIE 5163, 191-202, 2004.
13. F. Hoyle, and N. C. Wickramasinghe, “Comets-A Vehicle for Panspermia.” In Comets and the Origin of Life (C. Ponnaperuma, ed.), Reidel, Dordrecht, Holland, 227, 1980.
14. N. C. Wickramasinghe and F. Hoyle, “Infrared Radiation from Comet Hale-Bopp.” Astrophys. & Space Science 268, 379-381, 1999.
15. R. B. Hoover, F. Hoyle, N. C. Wickramasinghe, M. J. Hoover, and S. Al-Mufti, S., “Diatoms on Earth, Comets, Europa and in Interstellar Space.” Earth, Moon, and Planets, 35, 19-45, 1986.
16. E. V. Pikuta, and R. B. Hoover, “Psychrophiles and Astrobiology: Microbial Life of Frozen Worlds.” Instruments Methods and Missions for Astrobiology VI, SPIE 4939, 103-116, 2003.
17. C. F. Chyba, and C. Sagan, C., “Comets as a Source of Prebiotic Molecules for the Early Earth.” in Comets and the Origin and Evolution of Life. (Paul J. Thomas, Christopher F. Chyba, and Christopher P. McKay, Eds.) Springer-Verlag, New York. 147-168, 1997.
18. C. P. McKay, “Life in Comets.” in Comets and the Origin and Evolution of Life. (Paul J. Thomas, Christopher F. Chyba, and Christopher P. McKay, Eds.) Springer-Verlag, New York. 273-282, 1997.
19. W. F. Huebner, “Introduction” in Physics and Chemistry of Comets (Walter F. Huebner, Ed.) Springer-Verlag, New York, 2, 1990.
20. C. Emerich, J. M. Lamarre, V. I. Moroz, M. Combes, N. F. Sanko, Y. V. Nikolsky, F. Rocard, R. Gispert, N. Coron, J. P. Bibring, T. Encrenaz, and J. Crovisier, “Temperature and Size of the Nucleus of Comet P/Halley deduced from IKS infrared Vega 1 measurements.” Astron. Astrophys. 187, 839-842, 1987.
21. M. K. Wallis, and N. C. Wickramasinghe, “Structural Evolution of Cometary Surfaces,” Space Sci. Reviews 56, 93-97, 1991.
22. J. Klinger, “Classification of Cometary Orbits based on the concept of Orbital Mean Temperature,” Icarus, 55, 169-176, 1983.
23. A. H. Delsemme in Ices in the Solar System, (J. Klinger et al., Eds.) D Reidel, Dordrecht, Boston, Lancaster, 367-387, 1985.
24. M. K. Wallis, “Radiogenic Melting of Primordial Comet Interiors.” Nature 284, 431-432, 1980.
25. J. Klinger, “Composition and Structure of the Comet Nucleus and Its Evolution on a Periodic Orbit.” in Ices in the Solar System, (J. Klinger et al., Eds.) D Reidel, Dordrecht, Boston, Lancaster, 407-417, 1985.
26. M. J. S. Belton, W H Julian, A J Anderson, and B E A Mueller "The spin state and homogeneity of Comet Halley's nucleus", Icarus 93, 183-193, 1991.
27. D. Gilichinsky, E. Rivkina, V. Shcherbakova, K. Laurinavichuis and J. Tiedje, “Supercooled water brines within permafrost-an unknown ecological niche for microorganisms: a model for astrobiology.” Astrobiology, 3, 331-341, 2003.
28. H. U. Keller, “The Nucleus” in Physics and Chemistry of Comets (Walter F. Huebner, Ed.) Springer-Verlag, New York, 63, 1990.
29. K. Lodders and R. Osborne, “Perspectives on the comet-asteroid-meteorite link.” Space Sci. Rev. 90, 289-297, 1999.
30. D. P. Glavin, P. Ehrenfeund, O. Botta, G. Cooper, and J. L. Bada, “Amino Acid Chemistry as a Link between Small Solar System Bodies and Carbonaceous Chondrites. In Near Asteroid Sample Return Workshop 2000, http://www.lpi.usra.edu/meetings/asteroid2000/pdf/8003.pdf
31. M. H. Engel, V. E. Andrus, and S. A. Macko, “Amino Acids: Probes for Life’s Origin in the Solar System.” In “Perspectives in Astrobiology” (R. B. Hoover, R. Paepe and A. Yu. Rozanov, Eds.), NATO ASI, (In Press).
32. P. Ehrenfreund, D. P. Glavin, O. Botta, G. Cooper, and J. L. Bada, “Extraterrestrial amino acids in Orgueil and Ivuna: Tracing the parent body of CI type carbonaceous chondrites.” Proc. Nat. Acad. Sci., 98, 2138-2141, 2001.
33. NRC Report. http://planetaryprotection.nasa.gov/pp/bodies/minors.htm
34. R. B. Hoover, G. Jerman, A. Yu. Rozanov, and P. P. Sipiera, “New Evidence of Indigenous Microfossils in Carbonaceous Chondrites.” SPIE, 5555-2 (This Volume), 2004.
35. M. K. Wallis, and N. C. Wickramasinghe, "Interplanetary transfer of planetary microbiota" Mon. Not. Roy. Astron. Soc. 348, 52-61, 2004.